
¨BIOLOGÍA REPRODUCTIVA DEL TIBURÓN TORO (Carcharhinus leucas) Y SU POSIBLE USO DE ÁREAS DE CRIANZA EN TOPOLOBAMPO, SINALOA¨

1 Pelagios-Kakunjá A.C. Sinaloa 1540, Col. Las Garzas, C.P. 23070, La Paz, B.C.S., México.
2 College of Agricultural Sciences, Coastal Oregon Marine Experiment Station, Sulikowski Big Fish Lab, 430 Strand Agriculture Hall Corvallis, 97331, Oregon, USA.
3 Sealion Dive Center, Av. Gaviotas 7 Col. CETMAR, CP 81370, Topolobampo, Ahome, Sinaloa, México.
Palabras clave: Biología reproductiva, hormonas, áreas de crianza, ultrasonido, telemetría
INTRODUCCIÓN
Los tiburones son susceptibles a la sobrepesca debido a su bajo potencial reproductivo, reducido número de crías, largos periodos de gestación, crecimiento lento y madurez sexual tardía (Walker 1992, Castro 1993). Las limitantes biológicas características de los elasmobranquios y el fracaso histórico de sus pesquerías implican que en todos los casos su pesca debe ser gestionada cuidadosamente (Anderson, 1990). Además, factores como el incremento en el esfuerzo de pesca y la degradación de importantes áreas de crianza como hábitats costeros, estuarinos y de agua dulce
contribuyen a su decremento poblacional (Camhi et al. 1998, Stone et al. 1998). Las áreas de apareamiento y cría de tiburones se consideran habitats críticos de máxima importancia para mantener la continuidad del reclutamiento (Heupel et al. 2007).
A pesar de nuestra limitada comprensión sobre la ubicación y uso de áreas de crianza de tiburones, cualquier área que genere una agregación de especies probablemente tendrá implicaciones deproporcionadas para la gestión, ya que representa un hábitat esencial o aumenta el riesgo de explotación pesquera (Pratt & Carrier, 2001). Por esta razón, identificar sitios de crianza adicionales y comprender el uso de sitios ya conocidos es esencial. Recientemente se ha encontrado que el tiburón toro (Carcharhinus leucas) migra hacia el Farallón de San Ignacio (Fig. 1) en el área de Topolobampo, Sinaloa-México, entre julio y noviembre. Esta pequeña isla árida está compuesta de rocas escarpadas y dentadas (536 m de diámetro y 142 m de altura) y está rodeada de aguas profundas (>3,000 m), salvo por las rocas hacia el norte (Maluf, 1983). Este farallón es bien conocido por su alta biodiversidad y su ubicación en la zona de transición biogeográfica conocida como la “Brecha de Sinaloa” (Reyes-Bonilla et al., 2024).
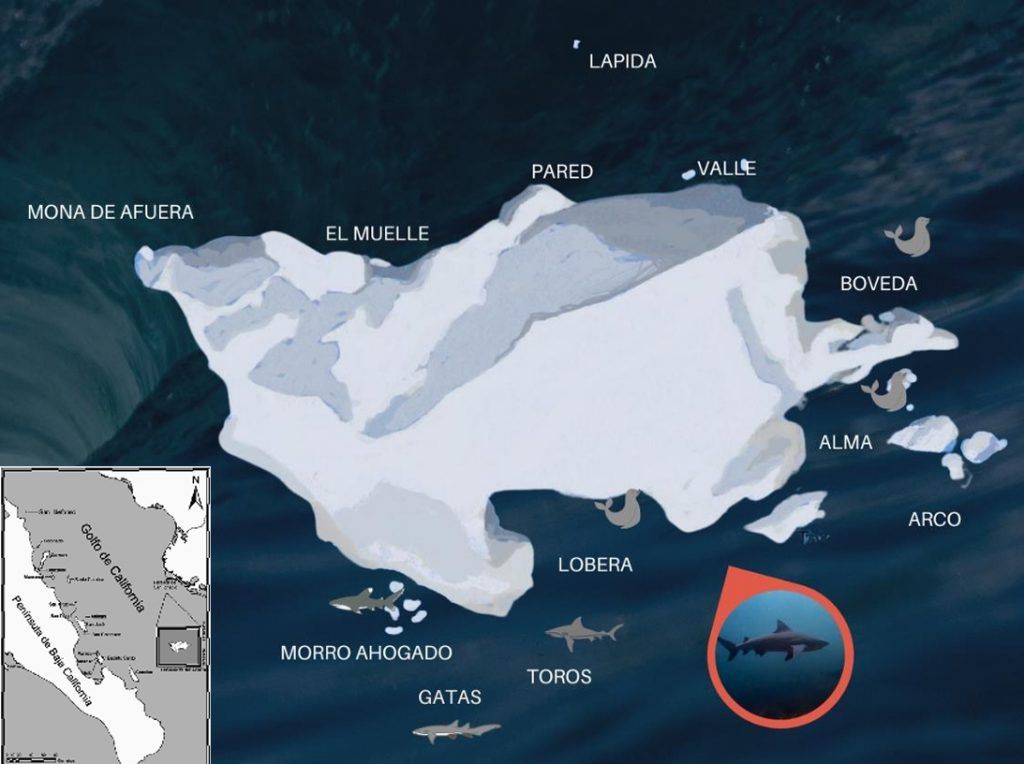
Aún no sabemos desde dónde migran los tiburones toro ni hacia donde, pero en cada inmersión durante la temporada se han registrado al menos seis ejemplares con longitudes totales que van desde 1.5 m LT (longitud total; juveniles) a 2.5 m LT (adultos). Un hecho interesante es que algunas de las hembras adultas han sido reportadas con marcas frescas de apareamiento durante el mes de septiembre (Fig. 2).

Los adultos se congregan para reproducirse en áreas de apareamiento, las cuales no necesariamente coinciden con las áreas de crianza (Hanchet 1988, Castro 1993). Comprender la influencia del comportamiento reproductivo en los movimientos de los peces y la selección de hábitat se ha reconocido como crucial para una gestión adecuada de los elasmobranquios. Aunque se ha documentado el uso repetido de áreas de crianza y gestación para varias especies de tiburones, se han identificado relativamente pocas áreas de apareamiento consistentes. Para la mayoría de las especies de tiburones, las áreas de apareamiento son desconocidas o se infieren a partir de los movimientos de los adultos, observaciones ocasionales de apareamiento en sitios de agregación anual o la presencia de heridas frescas de apareamiento observadas en momentos y lugares específicos (Pratt y Carrier, 2001).
Por otro lado, la presencia de juveniles en este farallón podría explicarse por la existencia de áreas de crianza cercanas. Éstas, son ubicaciones geográficamente discretas dentro del rango de distribución espacial de una especie, donde las hembras grávidas liberan a sus crías y donde los juveniles pasan sus primeras semanas, meses o años de vida (Castro 1993). Los hábitats de críanza de tiburón toro tienden a ubicarse en áreas con acceso limitado al océano y salinidad altamente variable, a menudo asociadas con islas de barrera y ensenadas estrechas (Bangley et al., 2018). El área de Topolobampo (38 km al este) comparte varias de estas características (Fig. 3). Los tiburones toro utilizan áreas de crianza caracterizadas por baja salinidad y a menudo, habitan ríos después de nacer (Snelson et al. 1984, Blackburn et al. 2007).
Se ha demostrado que esta especie tiene una preferencia por salinidades entre 7 y 20 PSU y se mueve activamente dentro de sistemas estuarinos a medida que cambian los flujos de agua dulce. Esta preferencia por la salinidad excluiría a la mayoría de las otras especies de tiburones y podría reducir sustancialmente el riesgo de depredación interespecífica (por parte de otras especies; Heupel & Simpfendorfer, 2008; 2011).

Aunque Topolobampo es una zona urbanizada, las bajas tasas de mortalidad en este tipo de áreas sugieren que el tiburón toro es capaz de adaptarse con relativa facilidad a ríos y estuarios urbanizados. Heupel & Simpfendorfer (2008) encontraron que las tasas de mortalidad natural de los tiburones toro neonatos en el río Caloosahatchee (Florida, EE. UU.) son más bajas en comparación con otras especies a pesar de ser un río altamente urbanizado, con desarrollos de canales, urbanización costera, descarga de aguas residuales tratadas, flujo de agua fuertemente gestionado y un tráfico pesado de embarcaciones. Se podría hipotetizar, por lo tanto, que los manglares y bahías poco profundas de Topolobampo podrían representar áreas de crianza para esta especie y que cuando los animales estén listos para dejarlas después de 4 años (Thorburn et al., 2008), se trasladan al Farallón de San Ignacio para alimentarse (Fig. 4).

Thorson (1976), trabajando con tiburones toro en Nicaragua, repotó que los tiburones toro neonatos (50-80 cm LT) se congregan en áreas de crianza de agua dulce y que los juveniles (80-100 cm LT) estaban casi completamente ausentes, sugiriendo que se encontraban en el mar. Snelson et al. (1984) encontraron que los juveniles (120 a 180 cm LT) están presentes en las aguas de la laguna durante todo el año, aparentemente por un período desconocido hasta que comienzan su maduración sexual.
Aunque la razón de la presencia de tiburones toro juveniles y adultos en este sitio permanece desconocida, Castro (1993) identificó otra área importante además de las áreas de crianza y apareamiento basándose en la distribución de tiburones por tamaño, sexo y sus patrones migratorios: las áreas de alimentación. El Farallón de San Ignacio alberga aproximadamente 500 lobos marinos de California (Zalophus californianus) y unos 700 lobos finos de Guadalupe (Arctocephalus townsendi; Fig. 1). En 2020, se registraron 492 lobos finos en enero y 771 en noviembre, la mayoría de los cuales eran juveniles (Gutiérrez‐Osuna et al., 2022). Estas dos especies podrían considerarse presas potenciales (especialmente los juveniles) para los tiburones toro en el Golfo de California, donde de hecho los pescadores locales los llaman “loberos” por alimentarse de estos animales.
Componente educativo comunitario
Los tiburones son animales majestuosos y tienden a recibir mucha atención de los medios de comunicación. Sin embargo, a menudo son estigmatizados. Pelagios Kakunjá está trabajando para cambiar esto proporcionando educación sobre tiburones y actividades de divulgación tanto para los niños en escuelas como para el público en general. Educar a la comunidad local conecta a las personas con el tiburón toro, una de las especies clave del ecosistema marino en el Golfo de California.
Nuestros programas no solo ayudan a proteger a los tiburones toro, sino que también brindan información que permite a las personas convivir en armonía con la naturaleza. Ofreceremos charlas sobre los tiburones toro para todos los niveles educativos en las escuelas locales de Topolobampo. Comenzando en octubre de 2024, nuestro Programa de Educación Ambiental alcanzó a unos 100 estudiantes del CETMAR (Centro de Estudios Tecnológicos del Mar; Fig. 5) a través de una charla informativa sobre la biología de los tiburones y los objetivos de nuestro proyecto. Para tener un mayor impacto social, colaboraremos activamente con otras instituciones educativas de la zona.

Objetivo general:
Proporcionar una descripción de la biología reproductiva de los tiburón toro que visitan el Farallón de San Ignacio y su uso potencial de áreas de crianza en Topolobampo, Sinaloa.
Objetivos específicos:
• Determinar el ciclo reproductivo de los tiburones toro del Farallón de San Ignacio mediante análisis hormonal.
• Identificar el estado de desarrollo de los embriones cuando las hembras de tiburón toro se acercan a las aguas costeras de Topolobampo.
• Precisar el momento y la ubicación de las áreas de parto y las áreas de crianza de los tiburones toro en Topolobampo.
• Determinar las rutas migratorias de los tiburones toro al partir del Farallón de San Ignacio mediante telemetría acústica.
MÉTODOS
Guía Foto-Identificación
Estudios previos han utilizado la identificación fotográfica de tiburones para ayudar a colectar datos sobre diferencias espacio-temporales en la proporción de sexos y patrones de residencia, así como sobre movimientos a corta y larga distancia (Strong et al., 1996; Klimley & Anderson, 1996; Bonfil, 2005). Estos estudios utilizaron características que incluían marcas distintivas en el borde posterior de las aletas dorsal y caudal, la presencia de manchas pigmentadas en la aleta dorsal, cicatrices en los flancos y estimaciones de la longitud total. En el caso de los tiburones toro, los buzos tomarán fotografías de ambos lados para la guía de foto-identificación. Dado que los patrones de pigmentación son muy similares en los tiburones toro, solo se considerarán características conspicuas como marcas de nacimiento, laceraciones, cicatrices y la presencia de objetos como anzuelos, etiquetas convencionales, etc. (Fig. 6).

Hormonas
La necesidad de responder preguntas básicas sobre la historia de vida de los tiburones está bien establecida, pero ha sido problemática debido al tamaño, la movilidad y el estado de sus poblaciones (Hammerschlag y Sulikowski 2011; Natanson et al., 2019). Entre las características de la historia de vida; la biología reproductiva, como el lugar y momento en que ocurre el nacimiento de los tiburones toro, el calendario de los ciclos estacionales o la duración de la gestación en esta área, siguen siendo en gran parte desconocidas.
Una estrategia utilizada para estudiar de manera no letal la biología reproductiva de los elasmobranquios es el análisis de la concentración de hormonas esteroides en piel y músculo (Sulikowski et al., 2006). Las hormonas reproductivas se correlacionan directamente con los cambios morfológicos en el tracto reproductivo, específicamente durante la maduración (Sulikowski et al., 2016) y durante los ciclos reproductivos (Sulikowski et al., 2006). La recolección de biopsias de piel y músculo se realizará mediante buceo autónomo y el uso de un arpón equipado con un dardo especial para biopsias (Fig. 7). Aproximadamente 2 gramos de tejido se recolectarán de la base de la aleta dorsal de cada tiburón. Las muestras de tejido muscular se utilizarán para extraer estradiol (E2), testosterona (T) y progesterona (P4), siguiendo los procedimientos de Prohaska et al. (2013). Los perfiles hormonales obtenidos permitirán la creación de una línea base de estas sustancias durante el ciclo reproductivo de los tiburones toro, relacionando los perfiles hormonales con el sexo, tamaño y mes.

Examinación con ultrasonido
Los tiburones toro serán capturados con anzuelo y línea por pescadores locales y muestreados en superficie. Los tiburones serán colocados al costado de la embarcación y asegurados con una cuerda en la cola, atada a una cornamusa en la popa y la línea con el anzuelo a una cornamusa en la proa. Asimismo, se colocará una cuerda en el vientre para evitar tensión en el anzuelo. Se realizará una ecografía a lo largo de la región abdominal de todas las hembras adultas para evaluar su etapa reproductiva (Sulikowski et al., 2016). Se utilizará un ultrasonido portátil Ibex EVO (E.I Medical Imaging, Colorado, EE. UU.) con un transductor curvado lineal de 60 mm y 5-2,5 MHz (modelo 290470), capaz de alcanzar una profundidad de escaneo de 24 cm, para obtener imágenes del tracto reproductivo de cada hembra. El escaneo se realizará en la superficie ventral, desde la aleta pectoral hasta la aleta pélvica para obtener imágenes transversales y longitudinales. Adicionalmente cada hembra será escaneada entre las aletas pectorales para observar los ovarios y los folículos. El aumento se ajustará automáticamente según la composición del tejido para optimizar lavisualización de los campos cercano y lejano en cada individuo. Las imágenes y secuencias de video recolectadas se utilizarán posteriormente para medir (a través del software propietario preinstalado en el ecógrafo Ibex EVO) el diámetro máximo del folículo (DMF) en milímetros y el diámetro de los embriones a lo largo del eje transversal en cm.
La sonografía se realizará en campo después de capturar a la hembra de tiburón toro y asegurarla en la embarcación con el vientre hacia arriba (Fig. 8). Además, se tomarán 5 ml de sangre de cada hembra a partir de la vena caudal para un análisis extra de hormonas reproductivas.

Las imágenes por ultrasonido tienen el potencial de ser utilizadas como una técnica no letal y recientemente, se ha vuelto más compacta y portátil. Aunque los estudios de tiburones mediante ultrasonido han sido limitados, esta técnica ha sido desarrollada por el Dr. Sulikowski, proporcionando un medio para evaluar visualmente la biología reproductiva y una determinación más precisa y no invasiva (Sulikowski y Hammerschlag, 2023).
Marca de Nacimiento (Birth tag)
Después de realizar la ecografía y en caso de embarazo, se insertará un dispositivo conocido como marca de nacimiento (Fig. 9). Esta marca (6 cm de largo por 2,5 cm de ancho) se insertará a través de la apertura cloacal, mediante un aplicador especializado, y permanecerá inactiva dentro del útero hasta el momento del parto. Dado el pequeño tamaño y forma de la marca, quedará oculta entre los embriones y una vez expulsada con las crías, flotará a superficie con el sensor húmedo-seco y la antena hacia arriba. El sensor se activa cuando detecta que la marca está seca (en superficie), momento en el que comienza a transmitir su ubicación mediante una señal a un satélite, que almacena los datos para su posterior descarga. Estas marcas se han utilizado exitosamente para determinar el momento y lugar de nacimiento del tiburón tigre (Galeocerdo cuvier) y del tiburón martillo común (Sphyrna lewini; Sulikowski y Hammerschlag, 2023). Estos procedimientos han sido aprobados por el Comité Institucional de Cuidado y Uso de Animales (IACUC) de la Universidad Estatal de Oregón.

TELEMETRÍA
Marcaje acústico. Se colocarán transmisores codificados (V16, Vemco Ltd., Halifax) de 71 mm de largo y 16 mm de diámetro, que emiten una señal de 69 kHz con una vida útil estimada de 10 años en los tiburones toro. Estos transmisores se colocarán externamente con un arpón en la base de la aleta dorsal del lado izquierdo mediante buceo autónomo. Además, los tiburones capturados para la ecografía serán marcados internamente dentro de la cavidad peritoneal mediante una pequeña incisión de 2.5 cm de ancho en la superficie ventral del cuerpo del tiburón (Fig. 10). La incisión se cerrará con tres suturas de material absorbible. El tiempo de manipulación se mantendrá por debajo de los 7 minutos.

Para estudiar la residencia y conectividad, se desplegará un conjunto de receptores automáticos (VR2W, Vemco Ltd., Halifax) con sensores de temperatura (Onset-HOBO) para detectar los transmisores codificados colocados en los tiburones toro. Cada vez que un tiburón marcado nade dentro del rango del receptor, éste registrará la identidad codificada del individuo junto con la fecha y hora de su paso. Los receptores se instalarán en el fondo del Farallón de San Ignacio con un anclaje compuesto de una cadena de acero galvanizado, línea de polipropileno y una boya de plástico (Fig. 11).
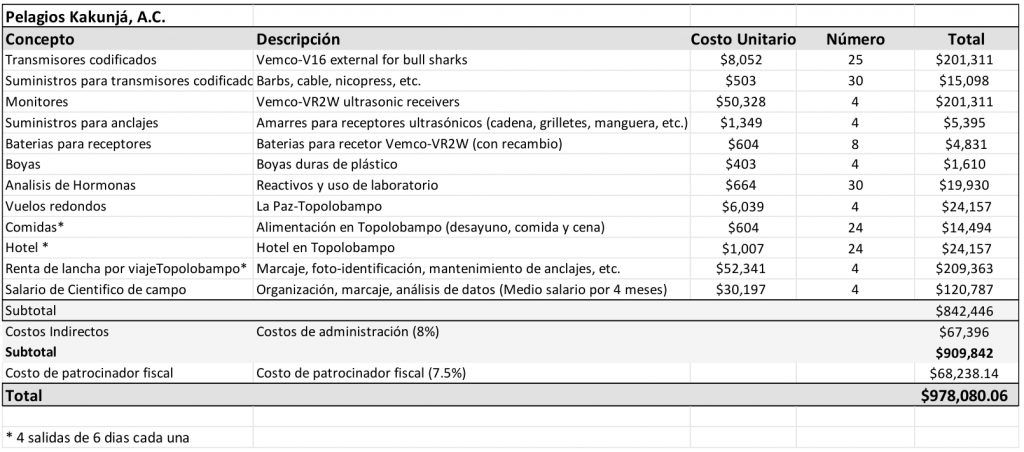
PRESUPUESTO (PESOS MEXICANOS)
REFERENCIAS
• Bangley, C. W., Paramore, L., Shiffman, D. S., & Rulifson, R. A. 2018. Increased abundance and nursery habitat use of the bull shark (Carcharhinus leucas) in response to a changing environment in a warm-temperate estuary. Scientific reports, 8(1), 1-10.
• Blackburn, J. K., Neer, J. A. & Thompson, B. A. 2007. In Shark nursery grounds of the Gulf of Mexico and the east coast waters of the United States. American Fisheries Society Symposium 50 (eds McCandless, C. T., Kohler, N. E. & Pratt, Jr., H. L.) 331–343 (American Fisheries Society, 2007).
• Camhi M., Fowler S., Musick J., Bräutigam A., Fordham S. 1998. Shark and their Relatives: Ecology and Conservation. IUCN Species Survival Commission, Occ. Pap. 20, 39 pp.
• Castro J. 1993. The shark nursery of Bulls Bay, South Carolina, with a review of the shark nurseries of the sourtheastern coast of the United States. Env. Biol. Fish. 38: 37–48.
• Gutiérrez‐Osuna, M. D. C., Díaz‐Gaxiola, J. M., Trites, A. W., & Hernández‐Camacho, C. J. 2022. New colony of Guadalupe fur seals (Arctocephalus townsendi) discovered on El Farallón de San Ignacio Island, Gulf of California. Marine Mammal Science, 38(1).
• Hammerschlag, N and J.A. Sulikowski. 2011. Killing for Conservation: Assessing the Need for Lethal Sampling of Apex-predatory Sharks. End. Spec. Res. Vol. 14: 135–140.
• Heithaus, M. R., and L. M. Dill. 2002. Food availability and tiger shark predation risk influence bottlenose dolphin habitat use. Ecology 83:480–491.
• Heupel, M., Carlson, J., Simpfendorfer, C. 2007. Shark nursery areas: Concepts, definition, characterization and assumptions. Mar. Ecol. Prog. Ser. 337: 287–297.
• Heupel, M. R. & Simpfendorfer, C. A. 2008. Movement and distribution of young bull sharks Carcharhinus leucas in a variable estuarine environment. Aquat. Biol. 1, 277–289.
• Heupel, M. R., & Simpfendorfer, C. A. 2011. Estuarine nursery areas provide a low-mortality environment for young bull sharks Carcharhinus leucas. Marine Ecology Progress Series, 433, 237-244.
• Lund R. 1990. Chondrichthyan life history styles as revealed by the 320 million years old Mississipian of Montana. Env. Biol. Fish. 27: 1–19.
• Maluf, L. Y. 1983. Physical oceanography. In T. J. Case & M. L. Cody (Eds.), Physical oceanography in the Sea of Cortez (pp. 26–45). University of California Press.
• Pratt, H.L., Carrier, J.C. 2001. A review of elasmobranch reproductive behavior with a case study on the nurse shark, Ginglymostoma cirratum. Environ Biol Fishes.; 60(1):157–188.
https://doi.org/10.1023/
• Prohaska, B.K., P. C. W. Tsang, W. B. Driggers III, E. R. Hoffmayer, Carolyn R. Wheeler, A. Christine Brown and J. A. Sulikowski. 2013. Assessing reproductive status in elasmobranch fishes using steroid hormones extracted from skeletal muscle tissue. Conser. Physiol. (1) 1-12.
• Reyes-Bonilla, H., Martínez-Castillo, V., Rojas-Montiel, B., & Olán González, M. 2024. Estructura comunitaria de peces, asteroideos y equinoideos en el Farallón de San Ignacio, arrecife situado en la “brecha faunística de Sinaloa”, México. Revista Ciencias Marinas y Costeras, 16(1), 01-19.
• Robbins W. D. 2006. Evaluation of two underwater biopsy probes for in situ collection of shark tissue samples. Mar. Ecol. Prog. Ser. 310: 213–217.
• Snelson, F. F., Mulligan, T. J., & Williams, S. E. 1984. Food habits, occurrence, and population structure of the bull shark, Carcharhinus leucas, in Florida coastal lagoons. Bulletin of Marine Science, 34(1), 71-80.
• Sulikowski, J.A., J. Kneebone, S. Elzey, W.H. Howell and P.C.W. Tsang. 2006. Using the composite parameters of reproductive morphology, histology and steroid hormones to determine age and size at sexual maturity for the thorny skate, Amblyraja radiata, in the western Gulf of Maine. J. Fish. Bio. 69: 1449-1465.
• Sulikowski, J.A., Wheeler, C.R. Gallagher, A.J., Prohaska, B.K., J.A., Langan and Hammershlag, N. 2016. Seasonal and life-stage variation in the reproductive ecology of a marine apex predator, the tiger shark (Galeocerdo cuvier), at a protected female aggregation site. J. Aqua. Biol. Vol. 24: 175– 184.
• Sulikowski, J. A., and Hammerschlag, N. 2023. A novel intrauterine satellite transmitter to identify parturition in large sharks. Sci. Adv. 9, eadd6340. doi: 10.1126/sciadv.add6340.
• Thorburn, D. C., & Rowland, A. J. (2008). Juvenile bull sharks’ Carcharhinus leucas'(Valenciennes, 1839) in northern Australian rivers. Beagle: Records of the Museums and Art Galleries of the Northern Territory, The, 24, 79-86.
• Thorson, T. B. 1976. The status of the Lake Nicaragua shark: an updated appraisal.
¿Y tú, te sumas?
Tu interés puede marcar la diferencia
Cada fotografía, cada inmersión, cada dato que recolectamos es un paso más hacia la conservación del tiburón toro y de todo el ecosistema marino que lo rodea. Pero este trabajo solo es posible con el apoyo de personas como tú, que creen en la ciencia, en la naturaleza y en el poder de actuar desde lo local.
Si esta historia te tocó, apóyanos.
Si te inspiró, compártela.
Y si te apasionó, únete a la causa.
"Ayúdanos a seguir investigando y protegiendo al tiburón toro."
Cada donativo se convierte en inmersiones, datos y conservación.